鸟类是适应飞翔的高等脊椎动物,具有极高的多样性和丰度。目前世界上鸟类的数量高达500亿只,在10000种鸟类物种中至少有40%为迁徙鸟类。为了利用夜间的有利天气条件和降低白天被捕食的风险,数十亿只通常在白天活动的小型雀形目鸟类转向夜间迁徙。尽管这些夜间迁徙的鸟类能够作为一类丰富的食物资源,但奇怪的是很少有捕食者能够捕食它们。
为什么能够捕食夜间迁徙鸟类的动物如此稀少?这是由于获取该资源需要捕食者克服行为、感官和生理上的众多挑战。首先,捕食者需要具有能够进食鸟类所需的头骨形态,因为头骨形态决定咬合力的大小,较大的咬合力才能撕咬和咀嚼鸟类食物。其次,由于夜间迁徙鸟类通常在高空以快速的飞行速度迁徙,捕食者需要具有空中飞行捕食的能力以及较快的飞行速度以追逐到鸟类。同时,捕食者需要具备能够在黑暗或光线较弱的条件下主动捕食鸟类猎物的感官能力。此外,高空和高速飞行意味着捕食者要承担快速而剧烈的能量消耗,因此捕食者需要适应为之而引起的一系列生理反应,例如低氧状况(需氧量不足)。最后,捕食鸟类后需要面临的一个重要挑战是对鸟类食物的消化、吸收和代谢,而在这一过程中,肠道微生物、甚至基因组层面上的变化起到关键作用,这就需要捕食者在这两个层面上做出适应性的改变和演化,以此来完成整个觅食鸟类的过程。因此,只有满足上述条件的动物才能征服这一具有挑战的觅食生态位。
迄今为止,仅发现毛翼山蝠(Nyctalus lasiopterus)、日本山蝠(Nyctalus aviator)和南蝠(Ia io)这3种神秘的蝙蝠能够捕食夜间迁徙的鸟类。然而,它们如何克服上述诸多挑战去捕食夜间迁徙鸟类,这对人们来说依然十分神秘。为了揭开蝙蝠捕食夜间迁徙鸟类的奥秘,东北师范大学动物生态与保护生物学研究团队以在中国分布的南蝠(图1)为研究模型,通过整合分子生物学、生理学、行为学、感官生态学、微生物学和基因组学的分析,以揭示南蝠如何克服行为、感官和生理上的一系列挑战去拓展这个极具挑战性的觅食生态位。

图1.南蝠
首先,研究人员采集蝙蝠粪便,通过分子技术(DNA宏条形码技术)分析和鉴定食性,发现南蝠在夏季主要捕食昆虫,在秋季主要捕食鸟类,并且捕食至少7科22种的小型雀形目鸟类(图2)。此外,绝大多数被捕食的鸟类具有迁徙习性,表明南蝠可能主要在飞行中采取空中飞行捕食策略捕食夜间迁徙的鸟类(论文1)
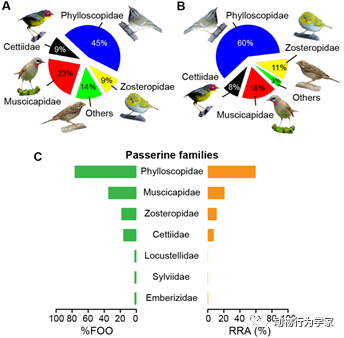
图2
南蝠食物中的鸟类猎物组成和多样性。(A)鸟类物种科水平出现百分比。(B)鸟类猎物条目科水平出现百分比。(C)鸟类猎物科水平出现频率百分比(%FOO)和相对序列丰度(RRA)。鸟类图片已被授权从鸟网(https://www.birdnet.cn/)下载、编辑和引用。
其次,研究人员选择与南蝠栖息在同一山洞和亲缘关系较近的13种不同食性蝙蝠为研究对象,共同构建系统发育树,并测量蝙蝠物种活体咬合力和扫描采集头骨三维形态数据,在系统发育背景下探究南蝠捕食鸟类的头骨形态和咬合力的相关性和适应性。结果发现不同食性蝙蝠的头骨形状和大小存在显著差异。南蝠具有较大的头骨尺寸和头部大小,拥有强健的下颌、缩短的齿列、较高的冠状突、扩张的角突、较低的髁突、较高的颅骨矢状嵴、缩短的眶间距、较宽的面部和颧弓(图3),这些高机械优势形态特征的演化使它们产生高咬合力来捕食、撕咬和咀嚼鸟类(论文2)。
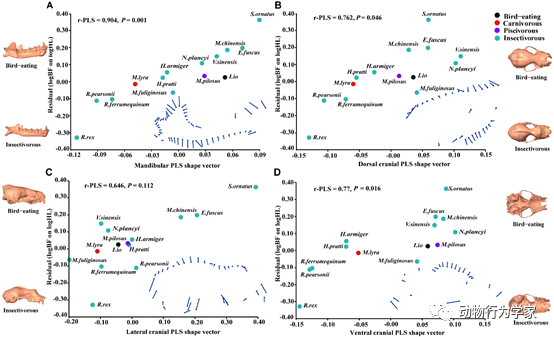
图3
南蝠捕食鸟类的头骨形态和咬合力适应。咬合力残差与头骨形状向量的偏最小二乘回归(PLS)分析。(A)下颌骨;(B)颅骨背面;(C)颅骨侧面;(D)颅骨腹面。右下方为形状变化,图左和图右为食鸟蝙蝠(上)与食虫蝙蝠(下)头骨。
此外,研究人员通过在野外给南蝠佩戴GPS设备,追踪和定位它们夜间捕食活动的空间位置。通过比较分析后发现南蝠在捕食迁徙鸟类季节(秋季)和食虫季节(夏季)呈现出不同的捕食空间活动模式。在秋季的捕食飞行海拔高度和距离地面飞行高度显著高于夏季,并且与小型雀形目鸟类的迁徙飞行高度一致。与此同时,记录到有3只个体在捕食鸟类的秋季飞行海拔高度超过了4500 m,最高为4945 m。此外,他们发现南蝠的飞行速度能够满足捕食夜间迁徙鸟类的速度需求。令人惊讶的是,南蝠在秋季的飞行速度可以达到143.7 km/h(图4)。这些结果揭示了南蝠能够克服捕食夜间迁徙鸟类的飞行空间高度与速度挑战(论文3)。
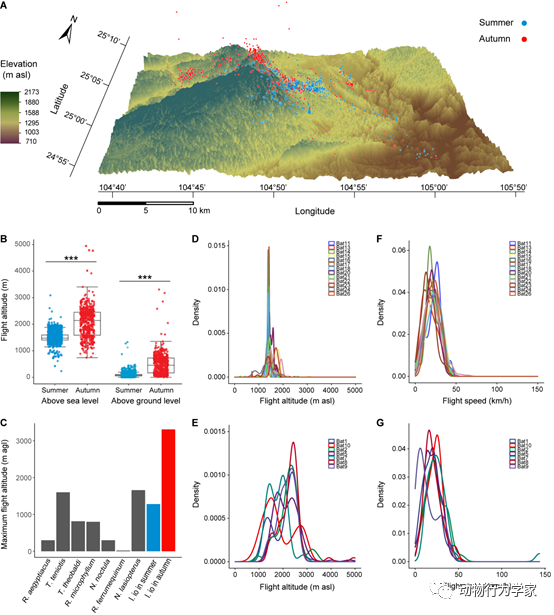
图4
南蝠捕食活动的飞行高度和飞行速度。(A)夏季和秋季所有个体的有效3D飞行位点数据。(B)夏季和秋季飞行高度的比较,分别以海平面和地平面作为参照。(C)利用GPS跟踪技术记录的7种蝙蝠物种地面以上的最大飞行高度以及南蝠夏秋季地面以上最大飞行高度。(D)夏季和(E)秋季南蝠个体飞行海拔高度核密度分布图。(F)夏季和(G)秋季南蝠个体飞行速度核密度分布图。
在食性研究的基础上,研究人员选取南蝠食谱中代表性的鸟类及其叫声和标本开展南蝠捕食鸟类的行为和感官模拟实验。捕食策略模拟实验发现,在黑暗条件下,南蝠在飞行中会主动攻击悬挂的鸟类标本,并且每一次攻击鸟类标本的过程中均发出完成一个空中飞行捕食猎物任务需要的声波序列,结束阶段具有捕食蜂鸣(feeding buzz)。捕食蜂鸣是回声定位蝙蝠捕食猎物之前才发出的,特点为脉冲间隔时间减少,重复率增加。因此,这一结果表明南蝠能够基于回声定位线索利用空中飞行捕食策略捕食鸟类。双选择声学回放结合基于听性脑干反应的听力测定实验发现,南蝠对回放其食谱中两种代表性鸟类(黄眉柳莺和暗绿绣眼鸟)的叫声与空白对照相比没有显著的选择偏好,且对回放频率低于6 kHz和声强低于70 dB peSPL的声音刺激不敏感。双选择视觉呈现实验结果显示,南蝠对暗光条件下的鸟类标本与对黑暗和月光条件下的鸟类标本的选择偏好次数也没有显著差异(图5)。这些结果共同表明,南蝠利用主动回声定位感官线索能够在夜空中飞行探测并定位捕食鸟类,不受限于鸟类是否发出叫声和环境光照强度的变化,突破了夜晚低可见度的视觉感官限制(论文3)。
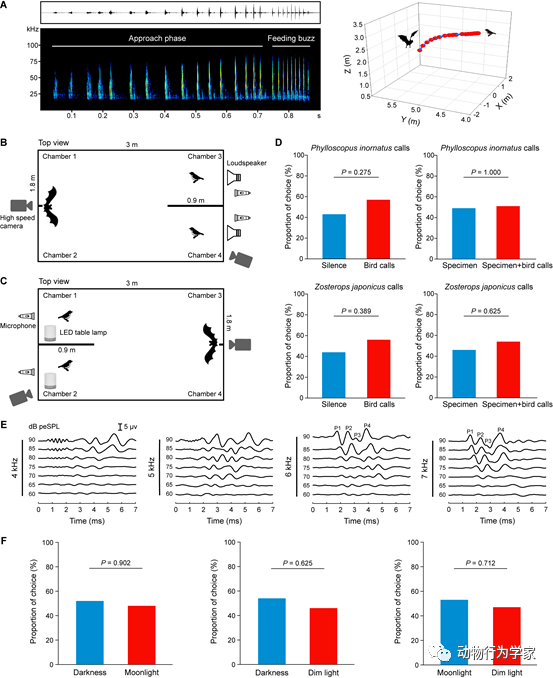
图5
南蝠空中飞行捕食鸟类及其捕食鸟类的感官线索。(A)南蝠攻击鸟类标本的发声序列及重建的三维飞行轨迹。(B)听觉线索实验装置俯视图。(C)视觉线索实验装置俯视图。(D)听觉线索实验中,空白与鸟叫声和标本与标本+鸟叫声实验组中的蝙蝠选择偏好次数百分比。(E)南蝠的低频听力(呈现4~7 kHz)。(F)视觉线索实验中,蝙蝠对呈现黑暗、月光和暗光不同光照强度实验组合中的蝙蝠选择偏好次数百分比。
最后,为了揭示南蝠在肠道微生物和基因组层面上可能发生的与捕食鸟类相应的适应性改变和进化。研究人员结合了肠道微生物、全基因组测序手段和比较基因组学的分析。结果发现,秋季捕食鸟类的南蝠肠道微生物群落结构组成和功能发生显著改变,与它们适应对鸟类食物中能量和营养的吸收利用,以及捕食鸟类过程中更高的能量消耗有关(图6)。此外,南蝠在DNA损伤修复、低氧适应、消化代谢和咀嚼肌发育方面的相关基因发生了适应性进化(图7)。这些结果说明,南蝠克服了高空飞行捕食鸟类所面临的低氧环境以及进食和消化代谢鸟类所要面对的生理挑战(论文3和论文4)。
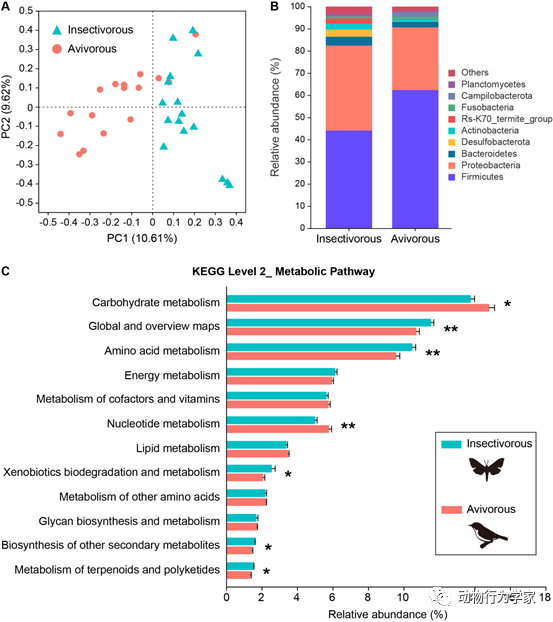
图6
南蝠捕食鸟类的肠道微生物适应。(A)基于Bray-Curtis距离的不同食性南蝠肠道微生物群落结构的主坐标分析图。(B)食虫和食鸟南蝠组肠道微生物在门水平的组成。“Others”表示其余所有细菌门的相对丰度之和。(C)食虫和食鸟蝙蝠组之间基于第二级KEGG通路的代谢相关类别的微生物宏基因组预测功能。
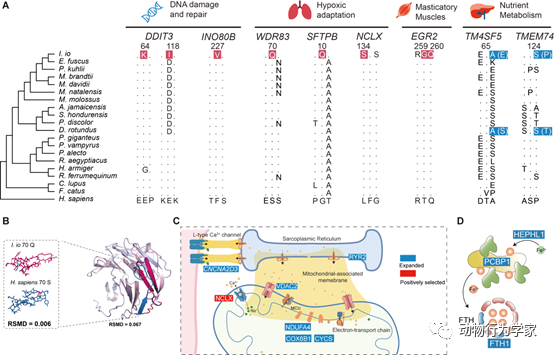
图7
南蝠捕食鸟类行为有关的基因适应性。(A)南蝠适应高空飞行的相关基因正选择分析结果,其中红色背景表示发生正选择的位点,蓝色表示趋同进化的位点。数字表示相应位点的位置,“.”表示该位点氨基酸和人类相同。(B)WDR83蛋白质三维结构预测,紫色表示南蝠WDR83蛋白质三维结构预测的结果,蓝色表示使用人类的氨基酸替换正选择位点的氨基酸后的三维结构。(C)在心肌细胞线粒体有关基因中的进化情况,红色表示正选择基因,蓝色表示扩张的基因。(D)南蝠铁离子代谢中进化的基因,蓝色表示扩张的基因。
这些工作系统、全面地揭示了南蝠捕食鸟类的头骨、行为、感官、肠道微生物和基因组适应,解开了蝙蝠能够捕食夜间迁徙飞行鸟类的奥秘。研究结果对深入理解食肉蝙蝠,尤其是食鸟蝙蝠的生态适应及其与猎物间的互作关系具有重要科学意义。东北师范大学的师资博士后龚立新、在读博士生耿炀、王梽强、林爱青副教授和硕士毕业生适碧叶为这些论文的第一作者或共同第一作者,冯江教授和江廷磊教授为论文共同通讯作者。这些研究得到国家自然科学基金项目(31922050, 31872680, 31872681, 32071492)和吉林省科技发展基金项目(20200201186JC)的资助。
论文1 链接:
https://onlinelibrary.wiley.com/doi/10.1002/ece3.7667
论文2链接:
https://frontiersinzoology.biomedcentral.com/articles/10.1186/s12983-020-00354-0
论文3 链接:
https://www.cell.com/iscience/fulltext/S2589-0042(22)01245-7
论文4 链接:
https://journals.asm.org/doi/10.1128/mSphere.00467-21
研究组网站:http://bat.nenu.edu.cn/




